

The innate cell recognition systems are due to surface or intracellular proteins in epithelial cells, neutrophils, macrophages and dendritic cells and to soluble cytoplasmic receptors. All these receptors are generally called PRRs (Pattern Recognition Receptors) and bind to molecules PAMPs (Pathogen-Associated Molecular Patterns) expressed only by the pathogen (lipopolysaccharide, bacterial carbohydrates and lipoproteins, peptidoglycans, RNA, DNA etc.) as well as commensal flora.

Pathogen recognition: innate immunity mechanisms
TLR (Toll-like receptors). Ten different members have been identified in pig. They are able to recognize bacterial products outside the cells and viral nucleic acid inside the cell. Thus, some TLRs are localized in intracellular compartments to recognize viral or bacterial nucleic acid. Other TLRs are on the cell surface and recognize bacterial products (lipoproteins, peptidoglycans, lipopolysaccharides).
Retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) are cytoplasmatic receptors recognizing viral dsRNA.
NOD (Nucleotide-binding oligomerization domain) proteins are cytosolic PRRs which cooperate with TLRs in recognizing bacterial products.
CLRs (C type-lectin receptors) is a surface receptors family including collectins, pentraxins, selectins, dectines and mannose receptor.
Proteins belonging to the complement system and to the acute phase proteins are able to act as a soluble receptor of recognition: C3 fragment, Mannose Binding Protein (MBP), surfactants, C Reactive Protein (CRP).
The mutual activation of cells at different tissue level, epithelial and endothelial cells and, especially inflammatory and innate cells, is the first physical and functional barrier against pathogens by triggering the inflammatory response.
Table 1: Systems of innate recognition of the pathogen.
| RECEPTORS | RECOGNITION ACTIVITY | ||
| TLR family (membrane or intracellular) | Recognition of bacterial and fungi products | TLR1 | Lipopeptides (bacteria and micoplasms) |
| TLR2 | Lipoproteins and proteoglicans (gram+) Lipopeptides (micoplasms) LPS (Leptospira) Lipoarabinomannan (mycobacteria) Lipopeptides (micoplasms) (gram+) Zimosan (fungi) |
||
| TLR4 | LPS (gram-) F-protein of RSV HSP60 (Chlamydia) |
||
| TLR5 | Flagellin (a lot of bacteria) | ||
| TLR6 | Regulates TLR2 Lipopeptides (micoplasms) Lipopeptides (micoplasms) (gram-) Zimosan (fungi) |
||
| TLR9 | CpG DNA of bacteria | ||
| TLR10 | Unknown | ||
| Recognition of viral products | TLR3 | dsRNA | |
| TLR7 TLR8 |
ssRNA. It’s restricted to pDCs and not expressed on cDCs | ||
| TLR9 | CpG DNA of virus. It’s restricted to pDCs and not expressed on cDCs | ||
|
CLRs |
Selectins, dectin 1,2 |
Recognizes carbohydrate domain on pathogen and induces phagocytosis or inflammosome activation, with pro-inflammatory cytokine production, in particular, addressing the immune response towards Th1 or Th17 finalization. | |
|
RLRs |
RIG-1 |
Binds ssRNA during the replication of viruses RNA, inducing the production of pro-inflammatory cytokines and type-I IFNs and conferring an antiviral state to the infected cells. | |
| NOD proteins (cytoplasmatic-intracellular) | NOD1 NOD2 |
Interaction with microbial (proteoglycan portions) components through their specific region. Porcine NODs are strongly expressed in mesenteric lymph nodes and in the gut associated lymphoid tissues (GALT), sustaining its role in the innate immune response against bacterial infections. | |
|
APPs (Acute Phase Proteins) |
C-Reactive Protein (CRP) | Produced in the liver, it binds phosphatidylcholine residues present on the wall of some bacteria and fungi, activating the complement system via classical manner and increasing phagocytosis by macrophages (opsonin-mediated phagocytosis). | |
| Mannose Binding Lectin (MBL) | It’s a collectin with lectin domains able to bind sugars containing mannose, so recognizing a wide number of viruses, bacteria, fungi and protozoa. MBL is able to activate the complement system in a classical or C1-independent manner which promotes opsonin-mediated phagocytosis. |
Pathogen recognition: specific immunity mechanisms
Specific B and T immune cells have receptors recognizing specific antigen determinants (epitopes) of the pathogen.
Naïve B lymphocytes have multiple surface receptor called BCR (B cell Receptor) constituted by a surface immunoglobulin, IgM or IgD, specific for an antigen only. During B cell development in bone marrow an enormous repertoire of BCRs, and consequently of B cells is created, by genetic recombination, to recognize an enormous number of specific antigens on the pathogens.
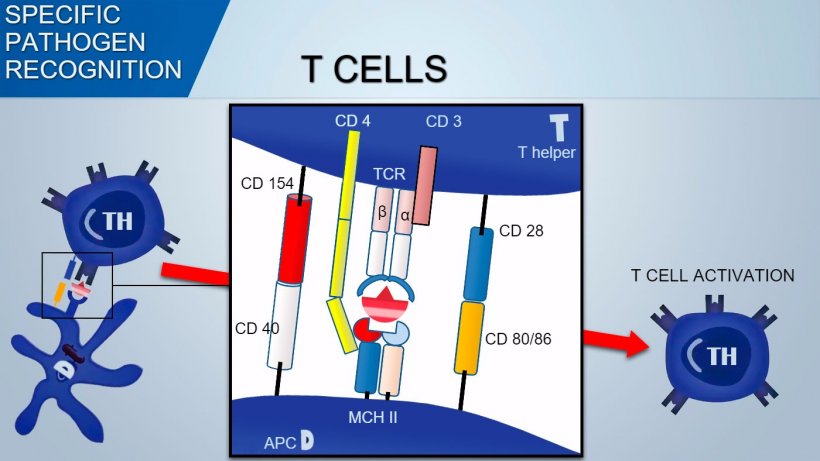
T Cell Receptor (TCR) of Thelper and cytotoxic T lymphocytes is constituted by two α and β chains. TCR of γδ T lymphocytes is constituted by two γ and δ chains. With a similar mechanism the variability of antigenic recognition of TCR on T-lymphocytes occurs.
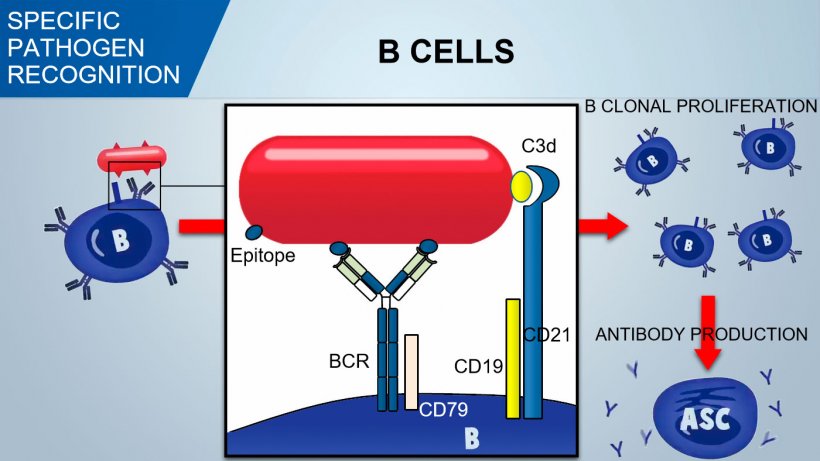
B cells recognize native antigenic determinants of the pathogen and the signal is transduced inside the cell by the activation of an associated glycoprotein (CD79α/β). Once internalized BCR-antigen complex, B cell process antigen and presents it in association with MHC II to Thelper lymphocyte. The co-stimulation between B and T cells is essential for B activation and to trigger an efficient antibody formation. An effective co-stimulation also requires a signal mediated by a CD21/CD19 complex which recognizes complement fragment (C3d) on pathogen.
Contrarily, T cell recognize antigen determinants processed and presented by APC in association with MHC (Major Histocompatibility Complex).
MHC II are molecules mostly present on the surface of professional APC such as mature dendritic cells, activated inflammatory macrophages and activated B cells. In turn, MHC I presents endogenous antigens to cytotoxic T lymphocytes.
MHC of pigs is known as SLA (Swine Leukocyte Antigen) and it’s constituted by 70 genes distributed in 3 classes:
In pigs, genes of MHC (SLA: “Swine Leucocyte Antigen”), are in the centromere of chromosome 7. Defined for the first time in 1970, seventy genes (haplotypes) are distributed in 3 classes:
- SLA I coding for MHC I molecules expressed on most nucleated cells other than neurons and trophoblasts. MHC molecules are associated and present antigens intracytoplasmic (eg. viral antigens) to CD8+ T cells. The MHC I molecules are constituted by one α heavy chain (highly polymorphic) and two β2 light chain called β2-microglobulin.
- SLA II: MHC class II molecules are expressed on B lymphocytes, on professional APC and on some subpopulations of T lymphocytes regardless of their activation status. MHC II molecules are associated with processed antigens (phagocytosed and degraded into peptides) inside the APC. MHC II molecules consist of α chain and β chain.
- SLA III are genes coding for the complement elements, cytokines etc.
To activate Th lymphocyte, signal mediated by TCR recognition of antigen-MHC complex should be associated with signal mediated by binding of surface co-stimulation molecules: CD28 (on T cell) binds to CD80 (on APC) or CD86 (on B cell), and CD40 (on APC) binds to CD154 (on T cell). This co-stimulation enhances the production of cytokines for T cell proliferation (IL-2) and also cytokine-drived Th differentiation.



