

Introduction
There is evidence that increasing feed intake in late gestation improves piglet birth weight (Cromwell et al., 1989, Shelton et al., 2009, Soto et al., 2011). However, the effect of increased amounts of feed offered during late gestation on piglet birth weight in high-performing herds (> 14.5 total piglets born/sow) is unclear. Further, because all studies reported increased feed intake, which increased both energy and amino acids (AA), it is unclear if the influences on piglet birth weight are due to dietary AA or energy content. Therefore, the objective of this study was to determine the effects of AA and energy intake during late gestation on piglet birth weight and reproductive performance of high-performing gilts and sows housed under commercial conditions.

Procedures
At day 90 of gestation, a total of 1,102 females (741 gilts and 361 sows) were housed in pens by parity group (P1 or P2+), blocked by weight within each pen, and each female was randomly assigned to dietary treatments within weight block. The average parity for P2+ (sows) after farrowing was 4.1 ± 1.9 cycles. Dietary treatments consisted of combinations of 2 SID AA intakes (10.7 or 20.0 g SID lys/day and other AA met or exceeded the NRC [2012] recommendations as a ratio to lys) and 2 energy intakes (4.50 or 6.75 Mcal/day intake of NE) in a 2 x 2 factorial arrangement. All other nutrients met or exceeded the NRC (2012) recommendations.
Results and discussion
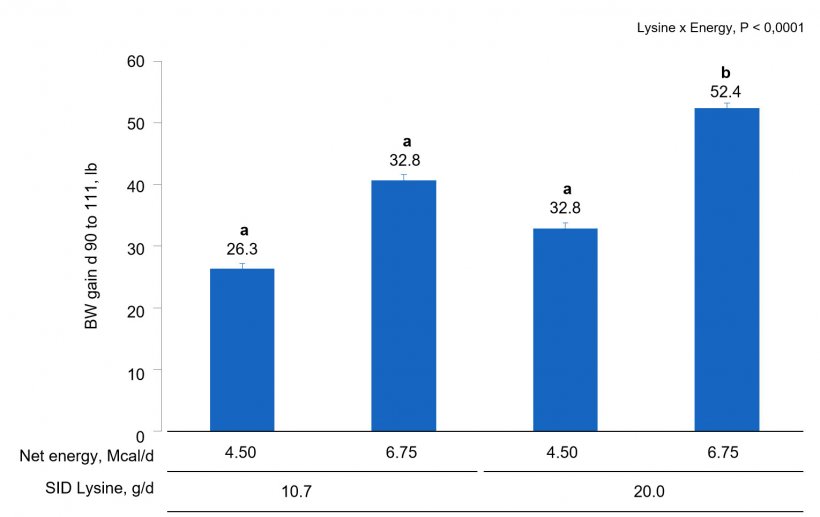
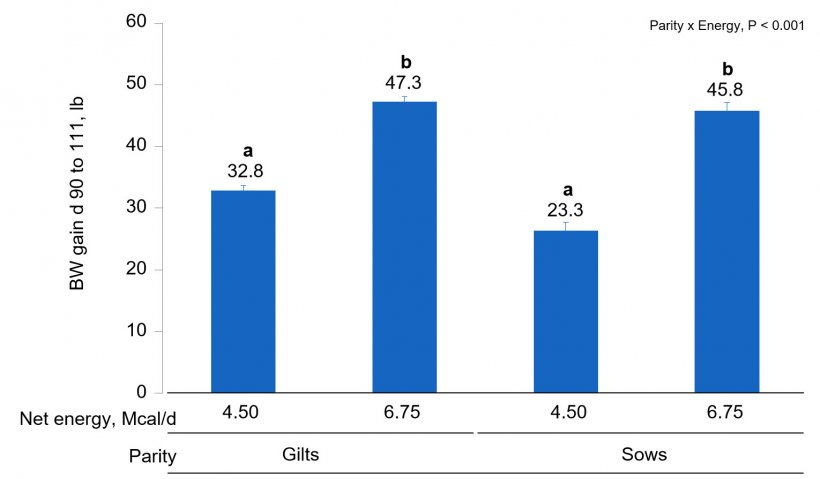
Chemical analysis of feed was similar to formulated values. In sows and gilts fed high-energy intake, the magnitude of body weight gain was greater with increasing AA intake (Fig. 1A) compared to increasing lys intake at low energy intake (AA × Energy, P < 0.001). Gilts gained more weight at low energy intake than sows (Fig. 1B; Parity × Energy, P < 0.001); however, there was no evidence for differences (P = 0.601) in weight gain between gilts and sows at high energy intake.
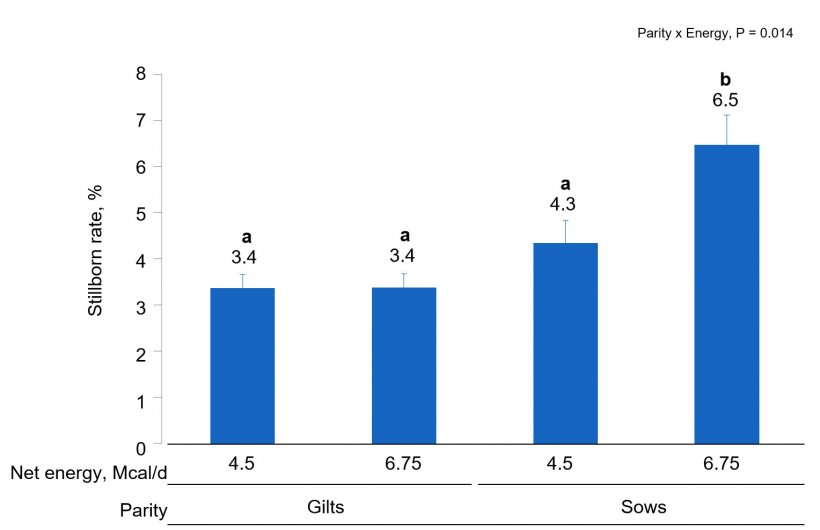
Sows had an increased number (P < 0.001) of total piglets born compared with gilts. Sows fed the high energy intake had marginally reduced probability of piglets born alive (Parity × Energy, P = 0.092) but there was no evidence for differences in gilts. This was due to the increased probability (Parity × Energy, P = 0.014) of stillborns in sows fed high energy intake (Fig. 2). The probability of stillborns was reduced (P = 0.049) in females fed high AA intake. There was a AA × Energy × Parity (P = 0.047) interaction where sows fed low energy and high AA intake had increased probability of mummified fetuses compared to sows fed low energy but low AA intake.
Considering the total number of piglets born, litter birth weight and individual piglet birth weight were heavier in sows (P < 0.001) than gilts, with marginally greater (P = 0.091) within litter birth weight CV in females fed high energy compared to low energy independent of parity. Within litter birth weight, CV was greater (P < 0.001) in sows than gilts. However, there was no evidence for differences between the dietary treatments for litter birth weight and individual piglet birth weight of total piglets born.
Individual born alive birth weight (Fig. 3) was heavier (P = 0.011) for females fed the high energy intake compared to low energy intake fed females, regardless of AA intake or parity. There was a lower percentage (P = 0.034) of pre-weaning mortality in piglets suckling from females fed high AA intake compared to low AA intake during late gestation regardless of energy level.
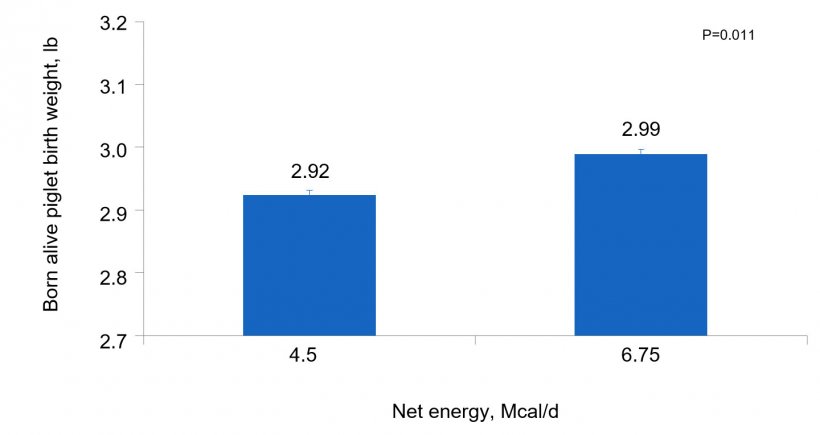
In conclusion, 1) body weight gain of gilts and sows depends not only on energy but also AA intake, 2) sows fed increased amount of energy had increased stillborn rate, 3) pre-weaning mortality was reduced in piglets suckling from females with high AA intake, and 4) the positive effect of increased amount of feed during late gestation on individual piglet birth weight, 30 g per pig, was due to energy rather than AA intake.

