
 Company
Boehringer Ingelheim Vetmedica GmbH
Germany
158
Publications
1066
Followers
Company
Boehringer Ingelheim Vetmedica GmbH
Germany
158
Publications
1066
Followers
Porcine Parvovirosis: an update
Porcine parvovirosis: an update
André Felipe Streck, Aline de Barros Moyses, Jéssica Martins Tadeu y Tamiris Silva Lopes
Laboratório de Diagnóstico em Medicina Veterinária, Universidade de Caxias do Sul
Abstract
Porcine parvovirus (PPV) is considered one of the main causes of reproductive disturbances in pig farming. Among the clinical signs commonly associated with PPV infection are fetal death, mummified fetuses, birth of stillbirths and delay in the return to estrus. Porcine parvovirus was always considered a conserved virus, without the constant emergence of new variants. However, there are suggestions that some parvoviruses exhibit a substitution rate close to some RNA viruses. In this context, new phenotypes of PPV have been reported in recent years, but its clinical importance and its capacity to reduce the efficacy of vaccines continues to raise questions. The aim of this review is to present an overview of the structure, pathogenesis and clinical aspects of PPV, highlighting the discovery of new profiles of capsids and studies on currently used vaccines.
In pig production, reproductive problems affecting herds cause large economic losses. Parvovirosis is one of the most common reproductive diseases that affects pigs. It is caused by porcine parvovirus (PPV) which has also recently been named Ungulate protoparvovirus 1 (Truyen and Streck, 2012). This virus was first isolated in Germany in 1965 as a contaminant of a primary swine cell culture used to propagate classical swine fever virus (Streck et al., 2015); since its discovery, PPV has remained a constant worldwide problem in that industrial sector (Mészáros et al., 2017). This viral infection is responsible for reproductive problems in females and mainly affects nulliparous swine. These disorders can be summarized with the acronym SMEDI (stillbirth, mummification, embryonic death, and infertility; Streck et al., 2015). At present, only one serotype has been identified; however, differences in the pathogenicity of different field strains have been reported and there are pathogenic and non-pathogenic strains (Truyen and Streck, 2012).
ETIOPATHOGENESIS
PPV is a very stable and highly infectious virus; both these factors facilitate spread of the infection and cause it to reach large parts of the swine population. Parvovirus can be introduced into herds through infected breeders and the virus can be transmitted via contaminated semen. In addition to semen, transmission can also occur through oronasal contact with infected animals or by contact with their secretions and excretions such as feces or fetal remains (Gradil et al., 1990; Streck et al., 2013). After infecting the animal, PPV replicates in their lymphoid tissues, salivary glands, lungs, bone marrow, and intestinal crypts (Duhamel et al., 1991). Because it replicates in intestinal cells, environmental contamination can occur which can cause the rapid spread of the virus (Carter and Saunders, 2013). In infected environments, the highest concentrations of PPV are found in farrowing rooms because PPV presents viral tropism towards embryonic and fetal tissues as well as to the fetal envelope (Truyen and Streck, 2012). Nulliparous females may be protected by the presence of maternal antibodies for up to six months of life, but this passive protection subsequently disappears at around the time sows begin their reproductive cycles (Truyen and Streck, 2012). If they are not immune to the virus, that is, if they are not vaccinated against it or if they were incorrectly vaccinated, pigs do not have any active immunity to PPV and will be more susceptible to infection. Consequently, they will be more likely to suffer from reproductive disorders (Carter and Saunders, 2013).
How does the infection evolve?
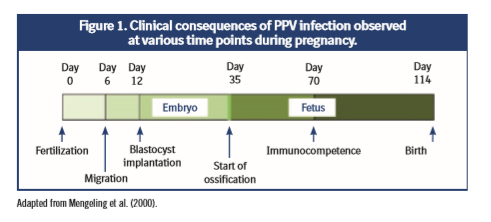
Vertical transmission to the fetus takes between 12 and 18 days after the mother is first infected via the natural (oral) route (Joo et al., 1976, Mengeling et al., 1980); this is because the virus can cause transplacental infection. The evolution of the infection depends on the sow’s stage of pregnancy at the time (figure 1):
- In the embryonic phase, until 30 days of gestation, the infection usually leads to death and reabsorption of the embryos. If most of the embryos die, the female may go back into heat at regular or irregular intervals; on the contrary, if most of the embryos resist the infection, the litter that is born will be small because the dead embryos will have been reabsorbed (Mengeling, 1979).
- From the 35th day of gestation, the fetal phase and ossification begins with the deposition of calcium in the fetal bones, which prevents reabsorption of the fetuses. When fetal infection occurs between 30 and 70 days of gestation, it is common for fetal death and mummification to occur. In general, when this happens the sow farrows a litter comprising both healthy and mummified piglets (Truyen and Streck, 2012).
- Immunocompetent fetuses infected after 70 days of gestation develop an antibody response and usually survive the infection. The virus is eliminated and antibodies against it will be detectable in the newborn’s serum (Truyen and Streck, 2012, Streck et al., 2015). One reason why PPV infection rates in the last third of gestation are low might be the lower rate of mitosis occurring in fetuses at this time which in turn, would reduce the viral replication rate (Mengeling et al., 2000).
CLINICAL MANIFESTATIONS
PPV infection is usually asymptomatic in adult animals. In most cases, the only evidence is reproductive insufficiency which manifests as repetitions, empty females, embryonic death followed by resorption, mummified fetuses, the birth of fewer, smaller, or weaker piglets, and neonatal mortality (Mengeling et al., 2000). Reproductive losses are usually low in vaccinated herds but PPV can cause a spate of abortions (a large number of abortions in a short period) in unvaccinated herds or in situations where the vaccine was incorrectly administered (Brown et al., 1980, Dea et al., 1985, Duhamel et al. 1991). Reproductive problems are mainly observed in nulliparous females (Mengeling et al., 1975, Mengeling et al.,1979). In multiparous females who have had previous contact with the agent, the action of the virus is reduced to zero (Truyen and Streck, 2012). In males, the infection is asymptomatic and sperm quality and libido are not affected (Christianson, 1992).
LESIONS
Mummification is usually observed in fetuses. Macroscopic lesions can include a varying degree of dwarfism before other external changes become evident (Figure 2).  Occasionally, blood vessels on the surface of the body become prominent as a result of their congestion and the filtration of blood through connective tissues. Congestion, edema, hydropericardium with hypertrophy of up to two ventricles, cerebral hemorrhage, congestion and hepatic necrosis, hyperemia and renal hemorrhage, hemorrhage with accumulation of sero-sanguinolent fluids in the body cavities, hemorrhagic discoloration that becomes progressively darker after death, and dehydration (mummification), are all typical of PPV infection. In addition, the placenta may be dehydrated and brown (Wilhelm et al., 2006). Because the virus is transmitted from one fetus to another during intrauterine infection, litter infection does not occur simultaneously. This explains the presence of mummified fetuses in different stages of fetal development, often alongside normal fetuses (Truyen and Streck, 2012). Microscopic fetal lesions tend to be generalized and are usually caused by cell necrosis in the developing organ systems (Lenghaus et al., 1978). Hemorrhages are often present in the subcutaneous tissues and muscle masses. Necrosis and mineralization are common in the lungs, kidneys, and skeletal muscle, and may be particularly extensive in the liver and heart (Lenghaus et al., 1978). If the infection occurs when the fetuses are immunocompetent, the microscopic lesions are mostly endometrial hypertrophy or mononuclear cell infiltration (Hogg et al., 1977). No macroscopic lesions are observed in females, but microscopically, it is possible to observe a mononuclear inflammatory infiltrate in the placental membranes and in the uterine epithelium, as well as the presence of lymphocytes in the central and medullary nervous system (Truyen and Streck, 2012).
Occasionally, blood vessels on the surface of the body become prominent as a result of their congestion and the filtration of blood through connective tissues. Congestion, edema, hydropericardium with hypertrophy of up to two ventricles, cerebral hemorrhage, congestion and hepatic necrosis, hyperemia and renal hemorrhage, hemorrhage with accumulation of sero-sanguinolent fluids in the body cavities, hemorrhagic discoloration that becomes progressively darker after death, and dehydration (mummification), are all typical of PPV infection. In addition, the placenta may be dehydrated and brown (Wilhelm et al., 2006). Because the virus is transmitted from one fetus to another during intrauterine infection, litter infection does not occur simultaneously. This explains the presence of mummified fetuses in different stages of fetal development, often alongside normal fetuses (Truyen and Streck, 2012). Microscopic fetal lesions tend to be generalized and are usually caused by cell necrosis in the developing organ systems (Lenghaus et al., 1978). Hemorrhages are often present in the subcutaneous tissues and muscle masses. Necrosis and mineralization are common in the lungs, kidneys, and skeletal muscle, and may be particularly extensive in the liver and heart (Lenghaus et al., 1978). If the infection occurs when the fetuses are immunocompetent, the microscopic lesions are mostly endometrial hypertrophy or mononuclear cell infiltration (Hogg et al., 1977). No macroscopic lesions are observed in females, but microscopically, it is possible to observe a mononuclear inflammatory infiltrate in the placental membranes and in the uterine epithelium, as well as the presence of lymphocytes in the central and medullary nervous system (Truyen and Streck, 2012).
PORCINE PARVOVIRUS GENETIC VARIATIONS
The genetic variations of PPV were not studied until the beginning of the 2000s. Until then it had been believed to be a very stable virus because vaccines developed from old strains still conferred immunity against variants of the virus (Truyen and Streck, 2012). With the discovery of new capsid profiles, the genetic variability of PPV has been studied over the last 20 years. In general, it was believed that viruses with DNA-based genomes were not very variable and so their study was not important in the control and prevention of diseases caused by PPV (Lopez-Bueno et al., 2006). This idea was initially formed because this type of virus uses the host’s cell polymerase complex which can maintain high-fidelity because of its error repair mechanisms (Drake, 1991). In contrast, viruses with RNA-based genomes, which do not have such error repair systems, have higher mutation rates (Lopez-Bueno et al., 2006). New studies revealed the existence of additional factors, such as oxidation, base methylation, and deamination, which can also affect mutation rates, even in DNAbased viruses (Duffy et al., 2008).
Emergence of new viral variants
The first study focused on the genetic variability of PPV strains obtained in the field in Brazil between 1994 and 2003 and was published in 2003 (Soares et al., 2003). In this study, 18 different capsid profiles were identified, and two phylogenetic groups were observed. Subsequently, another study analyzed samples isolated in 2001 and 2002 from Germany and described six different capsid profiles in another two phylogenetic groups (Zimmermann et al., 2006). After these studies, others were published that reported clinical events in different countries including Austria, China, Romania, and Switzerland (Shangjin et al., 2009; Cadar et al., 2011; Hao et al., 2011; Streck et al., 2011, Streck et al., 2013). In these studies, the strains with the highest prevalence in domestic pigs were related to strain 27a. Recently, an analysis of the viral capsid profiles found in domestic pigs showed that amino acid substitutions were mainly located in regions known as loops (figure 3; Truyen and Streck, 2012). Once the loops become located on the surface of the capsid, these amino acid substitutions can influence receptor binding and/or antigenic ability. Neutralizing antibodies play an important role in protecting against parvovirus infection and experimental infections with current vaccines have already shown that their protective activity is reduced against the predominant 27a strain (Zeeuw et al., 2007; Foerster et al., 2016).
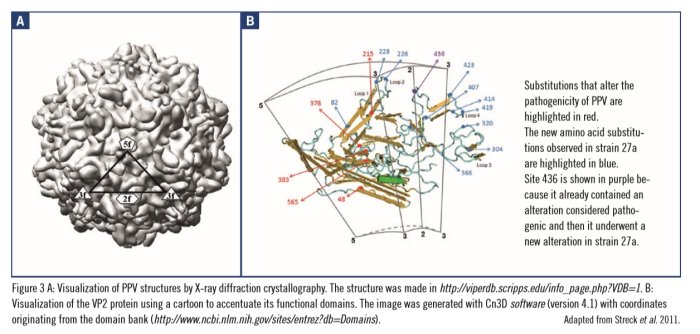
These results led to the hypothesis that the appearance of new amino acid substitutions may occur because of viral adaptation to the vaccines used and therefore, may represent ‘escape mutants‘ in a partially immune population. This possibility was studied by examining the dynamics of the viral population by using an in vitro and in silico model to reproduce possible immune selection scenarios (Streck et al., 2013). A moderate and continuous reduction in genetic diversity was observed from 1985. In other words, the worldwide use of PPV vaccines has probably not created these ‘escape mutants‘ but may have reduced PPV genetic diversity. This phenomenon can be observed when an agent that reduces or hinders the transmission of a virus is present (Van Ballegooijen et al., 2009) and therefore, some strains become predominant, with strain 27a standing out from among them (Streck et al., 2013).
VIRUS CONTROL
PPV is a virus that is relatively common in pigs and that is very resistant to the environment, which makes it difficult to control and prevent on farms (Truyen and Streck, 2012). Thus, vaccination, with the main objective of stimulating the immunity of these animals in order to avoid the intrauterine infection of embryos or fetuses, is the most common method so far used to control PPV (Days, 2010).
Vaccination
The vaccines currently available on the market are based on NADL-2 and include strains isolated more than 40 years ago. They are effective against homologous infections; however, they do not prevent infection or eliminate the virus after being exposed to challenges with antigenically heterologous strains. These vaccines are made with the inactivated virus (mostly with strains similar to Dusan the non-virulent NADL-2 strain; Truyen and Streck, 2012) and have a wide safety margin. In addition, they confer protection against PPV even if they are co-administered with other agents in polyvalent vaccines. The use of these vaccines induces good antibody titers which are sufficient to reduce clinical manifestations but unable to prevent PPV infection or spread (Jóźwik et al., 2009). However, modified live virus (MLV) vaccines may be an alternative for PPV. Other recombinant vaccines are being developed with more current technologies; however, inactivated vaccines are still the most common type used because they have such a wide safety margin (Truyen and Streck, 2012). The use of canine parvovirus MLV and feline panleukopenia virus induces a long-lasting immune response that protects pigs for several years. In addition, the occurrence and distribution of new PPV genetic variants must be ob
served very carefully. A new study not yet published observed that new mutations in capsid proteins can modify viral antigenic properties and reduce the binding and neutralization capacity of the antibodies generated by the commercial vaccines commonly used in pigs. It has also been proven that antibodies generated from predominant strains (27a) have a superior crossbinding capacity and can thereby protect pig populations more efficiently.
BIBLIOGRAPHY
- Brown TT, Paul Jr. PS y Mengeling WL.1980. Response of conventionally raised weanling pigs to experimental infection with a virulent strain of porcine parvovirus. American Journal of Veterinary Research, 41: 1221–1224.
- Cadar D, Dán Á, Tombácz K, Lőrincz M, Kiss T, Becskei Z, Spînu M, Tuboly T y Cságola A. 2012. Phylogeny and evolutionary genetics of porcine parvovirus in wild boars. Infection, Genetics and Evolution, 12: 1163-1171.
- Carter JB y Saunders VA. 2013. Parvoviruses (and Other ssDNA Viruses). Virology: Principles and Applications. 2. ed. West Sussex, Uk: Wiley. Cap. 12. p. 135-141.
- Christianson WT. 1992. Stillbirths, Mummies, Abortions and Early Embryonic Death. Veterinary Clinics of North America Food Animal Practice. Cap.8, p. 623639.
- Dea S, Elazhary MASY, Martineau GP y Vaillancourt J. 1985. Diseases of swine. Canadian Journal of Comparative Medicine, 49:343–345.
- Duffy S, Shackelton L y Holmes EC. 2008. Rates of evolutionary change in viruses: patterns and determinants. Nature Review Genetics, 9:267-276.
- Duhamel GE, Bargar TW, Schmitt BJ, Molitor TW y LU W. 1991. Identification of parvovirus-like virus particles in intestinal crypt epithelial cells of pigs with diarrhea. Journal of Veterinary Diagnostic Investigation, 3:96-98.
- Foerster T, Streck AF, Speck S, Selbitz HJ, Lindner T y Truyen U. 2016. An inactivated whole-virus porcine parvovirus vaccine protects pigs against disease but does not prevent virus shedding even after homologous virus challenge. Journal of General Virology, 97:1-6.
- Gradil C, Molitor T, Harding M y Crabo B. 1990. Excretion of porcine parvovirus through the genital tract of boars. American Journal of Veterinary Research, 359362.
- Hao X, Lu Z, Sun P, Fu UY, Cao Y, LI P, Bai X, Bao H, Xie B, Chen Y, Li D y Liu Z. 2011. Phylogenetic analysis of porcine parvoviruses from swine samples in China. Virology Journal, 8:320.
- Hogg GG, Lenghaus C y Forman AJ. 1977. Experimental porcine parvovirus infection of foetal pigs resulting in abortion, histological lesions and antibody formation. Journal of Comparative Pathology, 87: 539–549.
- Joo H, Donaldson-Wood C y Johnson R. Observations on the pathogenesis of porcine parvovirus infection. 1976. Archives of Virology, 51:123-129.
- Jóźwik A, Manteufel J, Selbitz HJ y Truyen U. 2009. Vaccination against porcine parvovirus protects against disease, but does not prevent infection and virus shedding after challenge infection with a heterologous virus strain. Journal of General Virology, 2437-2441.
- Lenghaus C, Forman AJ y Hale CJ. 1978. Experimental infection of 35, 50 and 60 day old pig foetuses with porcine parvovirus. Australian Veterinary Journal, 418.
- Lopez-Bueno A, Villarreal LP y Almendral JM. 2006. Parvovirus variation for disease: a difference with RNA viruses. Current Topics in Microbiology and Immunology, 299:349–370.
- Mengeling WL. 1979. Prenatal infection following maternal exposure to porcine parvovirus on either the seventh or fourteenth day of gestation. Canadian Journal of Comparative Medicine, 106.
- Mengeling WL, Paul PS y Brown TT. 1980. Transplacental infection and embryonic death following maternal exposure to porcine parvovirus near the time of conception. Archives of Virology, 65:55-62.
- Mengeling WL, Lager KM y Vordwald AC. 2000. The effect of porcine parvovirus and porcine reproductive and respiratory syndrome virus on porcine reproductive performance. Animal Reproduction Science, 199–210.
- Mészáros I, Olasz F, Cságola A, Tijssen P y Zádori Z. 2017. Biology of Porcine Parvovirus (Ungulate parvovirus 1). Viruses, 9: 393.
- Shangjin C, Cortey M y Segalés J. 2009. Phylogeny and evolution of the NS1 and VP1/ VP2 gene sequences from porcine parvovirus. Virus Research, 209–215.
- Soares RM, Cortez A, Heinemann MB, Sakamoto SM, Martins VG, Bacci M, De Campos FM y Richtzenhain LJ. 2003. Genetic variability of porcine parvovirus isolates revealed by analysis of partial sequences of the structural coding gene VP2. Journal of General Virology, 1505-1515.
- Streck AF, Bonatto SL, Homeier T, Souza CK, Gonçalves KR, Gava D, Canal CW y Truyen U. 2011. High rate of viral evolution in the capsid protein of porcine parvovirus. Journal of General Virology, 92:2628-2636.
- Streck AF, Homeier T, Foerster T, Fischer S y Truyen U. 2013. Analysis of porcine parvoviruses in tonsils and hearts from healthy pigs reveals high prevalence and genetic diversity in Germany. Archives of Virology, 158:1173-1780.
- Streck AF, Canal CW y Uwe T. 2015. Molecular epidemiology and evolution of porcine parvoviruses. Infection, Genetics And Evolution, 36:300-306.
- Truyen U y Streck AF. 2012. Porcine Parvovirus. In: Zimmerman, Jeffrey J. et al. Diseases of Swine. 10. ed. Uk: Wiley-blackwell. Cap. 29. p. 447-454.
- Van Ballegooijen WM, Van Houdt R, Bruisten SM, Boot HJ, Coutinho RA y Wallinga J. 2009. Molecular sequence data of hepatitis B virus and genetic diversity after vaccination. American Journal of Epidemiology, 170:1455-1463.
- Wilhelm S, Zimmermann P, Selbitz HJ y Truyen U. 2006. Real-time PCR protocol for the detection of porcine parvovirus in field samples. Journal of Virological Methods, 134: 257-260.
- Wrathall AE, Wells DE, Cartwright SF y Frerichs GN. 1984. An Inactivated, Oil Emulsion Vaccine For The Prevention Of Porcine Parvovirus-Induced Reproductive Failure. Research In Veterinary Science, 36:136-143.
- Zeeuw EJL, Leinecker N, Herwig V, Selbitz HJ y Truyen U. 2007. Study Of The Virulence And Cross-Neutralization Capability Of Recent Porcine Parvovirus Field Isolates And Vaccine Viruses In Experimentally Infected Pregnant Gilts. Journal of General Virology, 88:420-427.
- Zimmermann P, Ritzmann M, Selbitz HJ, Heinritzi K y Truyen U. 2006. VP1 Sequences Of German Porcine Parvovirus Isolates Define Two Genetic Lineages. Journal of General Virology, 87:295-301.
Contact:
Contact us using the following form.