

During the first days of life, piglets receive both bacteria and protective calostral antibodies from their mothers. This leads to the animals being colonised by H. parasuis while they are protected by the maternal immunity, until they are capable of producing their own immune response. The result is a balance between colonisation and antibodies that keeps the strains under control and prevents the development of disease. The decrease in the contact time between the mother and her piglets, due to the common practice of early weaning, has led to the appearance of some piglets that are not completely colonised or that have not acquired a sufficient level of maternal antibodies. On mixing with other litters, these piglets are faced with distinct strains while lacking the necessary antibodies against the disease.
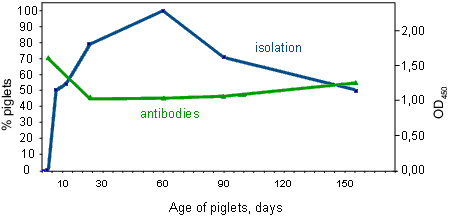
Fig. 1. In the first days of life, piglets receive immunity and bacteria from their mothers. Later, they produce their own response. In blue, percentage of piglets from which Haemophilus parasuis has been isolated. In green, antibodies (IgGs) in serum from the same animals.
The strains of H. parasuis can be classified into groups according to their serotype (see chapter “Epidemiology and typing”). Fifteen serotypes of H. parasuis have been defined, but there also exist a considerable quantity of strains that do not belong to any of these serotypes. When the 15 serotypes were defined, experimental infections were made with the reference strain of each serotype and a great variability of virulence was observed between them. Since then, it has been assumed that all the strains belonging to one serotype have the same degree of virulence as the corresponding reference strain. In some cases it seems likely that this association is real, as in the case of the strains of serotype 5. Serotype 5 was defined as being a highly virulent serotype, and it is also one of the most prevalent since due to its virulence the strains belonging to this serotype are frequently isolated from animals with Glässer’s disease. It is also one of the serotypes that has been used most in experimental infections, and this has led to the confirmation of its high virulence.
However, there are other data that are more contentious. In 1993, Neilson carried out studies on immunity and cross-protection, observing that after intranasal inoculation of live strains of the serotypes 1, 2, 3, 4, 5, 6 and 7, antibodies were produced with homologous and heterologous cross-reaction (see Table). Furthermore, the animals inoculated with serotypes 2, 3, 4 or 7 proved to be protected against an intranasal infection with a strain of serotype 5. These results indicate that the intranasal infection produces circulating antibodies to common antigens of distinct serotypes. On the contrary, in 2001 Takahashi et al observed that a bacterium of serotype 2 did not protect against an intratracheal infection with serotype 5, and vice versa. On the other hand, in 1997 Rapp-Gabrielson et al found that the intramuscular immunization with bacteria of distinct serotypes, followed by intratracheal infection, resulted in variable protection. It is worth noting the heterologous protection of a bacterium combined with serotype 4 and 5 against serotypes 13 and 14, but not serotypes 2 or 12, and more surprisingly the lack of protection of a serotype 12 bacterium against a different strain from the same serotype 12.
Table. Titles of antibodies of piglets inoculated intranasally with reference strains of distinct serotypes of Haemophilus parasuis.
| Serotype antigen | |||||||
| Inoculated serotype | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| 1 | 64 | 0 |
16 |
0 |
0 |
0 |
0 |
| 2 | 0 | 16 |
0 |
0 |
0 |
0 |
0 |
| 3 | 32 | 0 |
16 |
0 |
0 |
0 |
0 |
| 4 | 0 | 16 |
0 |
16 |
8 |
0 |
16 |
| 5 | 0 | 16 |
0 |
8 |
16 |
16 |
8 |
| 6 | 0 | 8 |
0 | 8 | 8 | 16 | 8 |
| 7 | 0 | 16 | 0 | 8 | 8 | 8 | 16 |
| Nielsen, R. Acta Vet. Scand. 1993, 34:193-198 | |||||||
The apparent contradictions in the literature could be due to the use of different inoculation routes, both for immunization and for the posterior infection, and to the use of distinct strains within the same serotype, since a variation in virulence has also been observed within the strains of the same serotype. The serotype is defined by the immunologic properties of the soluble antigen of the strains (possibly LOS and polysaccharides), but the specific composition of this antigen is unknown. The specific knowledge of the molecules that determine the serotype could help to clarify the cross-reactions between the strains of H. parasuis, which would be useful for establishing vaccination strategies.
